Stoneflies - Plecoptera: Perlodidae of Gunnison County, ColoradoMegarcys signata Laramide Springfly(Hagen 1874)
Updated 24 March 2024
TSN 103111

Description
Late instar Megarcys nymphs are easy to identify in the field because they´re big, active stonefly nymphs with small stubs of gills on each side of their thorax and neck. Look for the "W" on their heads as well. They become darker as they approach the time to emerge as adults.
Habitat
Under rocks in the swift portions of streams and rivers throughout Gunnison county. Present across the Coastal, Cascade and Rocky Mountains of the western United States.
Local Research Results
Megarcys signata nymphs have been popular research subjects in the upper Gunnison Basin around the Rocky Mountain Biological Lab (RMBL). Common and easy to catch in the summer, these stoneflies are robust and relatively easy to handle and observe (as are Kogotus modestus). Both the Peckarsky and Allan Labs at RMBL have worked with this species. In fact Megarcys is such a popular research subject, you may want to read the annotations in the references of this webpage to enjoy them :-)
 This adult was running around the rocks and spray in a steep tributary to Copper Creek on 18 July 2007.
This adult was running around the rocks and spray in a steep tributary to Copper Creek on 18 July 2007.Locations Collected
Copper Creek, East River, Benthette Brook, Cement Creek, Snodgrass Creek, Oh be Joyful Creek, Lake Fork Gunnison River and tributaries of all these streams. This author has specimens from the East River and Copper Creek near Gothic and Paradise Basin.
Notes
Hagen described this species in 1874 as Dictyopteryx signata, then it was called Perlodes signata, then renamed Arcynopteryx signata and now its called Megarcys signata. These generic name changes are common for many species as additional collecting reveals more information about the diversity or ranges of a group of insects. Some name changes occur when entomologists name species based on adults, then later manage to rear and associate nymphs and adults. Entomologists often discover traits of the nymphs that either match or are strikingly different from other genera. This causes everyone paying attention to the details to agree that the animal should be assigned to a different genera. At least the specific name (signata) stayed exactly the same through all this!
Good Links
On this website:
Photo - of Megarcys chasing an Epeorus nymph. You may need to page down once to see the photo.
Other Websites:
Photo - of Megarcys sp. nymph from the Tree of Life
http://tolweb.org/tree/eukaryotes/animals/arthropoda/hexapoda/plecoptera/slideshows/pleco1/sld031.htm

This adult was crawling around on a rock near a stream in an alpine basin near Crested Butte on the 19th of July 2007.
References
Allan, JD 1982 Feeding habits and prey consumption of three setipalpian stoneflies (Plecoptera) in a mountain stream. Ecology 63 1, 26-34. Abstract
Allan found that Megarcys has a 1 year life cycle in Cement Creek and is present above 2900m Early instar nymphs appear by the beginning of June, grow rapidly throughout the summer, overwinter as larvae and emerge the following late June, early July. Growth was rapid until the water temperature dropped to 0°C in mid-October. Final instar females weighed 25-40 mg and males weighed 12-17 mg. M. signata guts were found to be fullest at 0400 and 0800 when animals were collected every 4 hours over a 24 hour period. Further research showed that gut clearance times were longer when the water was colder at night. Therefore, taking into account the 1 day time periods for gut clearance in starved animals, it appears that M. signata feeds continuously and not primarily at night (Allan, 1982). M. signata fed primarily on diatoms and chironomids over the 24 hours period they were examined.
Allan,JD; Flecker,AS and McClintock,NL 1987 Prey size selection by carnivorous stoneflies. Limnology and Oceanography 32 4, 864-872.
Size selectivity varied with size of Megarcys. Percent of attacks per encounter by small stoneflies was strongly biased towards small prey. Large Megarcys attacks were weakly biased towards large prey. Capture success was greater and handling times were shorter with small prey compared to large prey.
Baumann,RW; Gaufin,AR and Surdick,RF 1977 The stoneflies (Plecoptera) of the Rocky Mountains. Memoirs of the American Entomological Society 31, 1-208. PDF
Quoted from page 131: "This species is common in creeks and rivers. The adults emerge from April to July"
Cather,MR and Gaufin,AR 1975 Life history and ecology of Megarcys signata (Plecoptera: Perlodidae), Mill Creek, Wasatch Mountains, Utah. Great Basin Naturalist 35 (1) PDF
Abstract:
"During investigation of some of the stoneflies of Mill Creek, Wasatch Mountains, Utah, Megarcys signata, a large omnivourous stonefly was found to have a univoltine life history and a slow seasonal life cycle.
Temperature appears to affect the growth rate of Megarcys signata. Warmer stream temperatures accompany the acceleration of growth rate, whereas cooler stream temperatures apparently retard the growth rate.
Periods of maximum absolute growth rate correspond with maximum carnivorous feeding from August to September and March to April. Chironomidae, Ephemeroptera and Plecoptera, in that order were the most abundant prey in the foreguts. Young nymphs ingested considerable amounts of diatoms, filamentous algae, and detritus but not as much animal matter as did older nymphs.
Megarcys signata was uniformly distributed throughout Mill Creek,except at the lowest station, where few nymphs were found.
Emergence occurred in May and June, the peak occurring in June. The mean size of females and males decreased as emergence progressed."
Colburn,T 1982 Measurement of low levels of molybdenum in the environment by using aquatic insects. 29, 422-428.
Gill,BA; Harrington,RA; Kondratieff,BC; Zamudio,KR; Poff,NL and Funk,WC 2014 Morphological taxonomy, DNA barcoding, and species diversity in southern Rocky Mountain headwater streams. Freshwater Science 33(1) 288-301.
Working in wadeable streams on the Front Range of Colorado, they found a cryptic species of Megarcys alongside M. signata.
Hagen,HA, 1874 Report on the Pseudo-neuroptera and Neuroptera collected by Lieut. W.L. Carpenter in 1873 in Colorado. Annual Report of the U.S. Geological and Geographical Survey of the Territories, embracing Colorado, 7: 571-577.
Described as Dictyopteryx signata.


Kjer,KM 2004 Aligned 18S and insect phylogeny. Systematic biology, 53(3), pp.506-514. PDF
In FIGURE 1. Phylogeny of Insecta from 18S rRNA data, it appears the author has a typo. M. signata is written as Megarcys stigmata.
Klapálek, Frantisek 1912 Plécoptères. I. Fam. Perlodidae; [monographische Revision. II. Fam. Perlidae; Subfam. Perlinae, Subfam. Neoperlinae; mongraphische Revision] Series Sélys-Longchamps, Edmond de, baron, 1813-1900. Collections zoologiques; catalogue systematique et descriptif, fasc. 4, pt. 1-2.

Knight,AW; Gaufin,AR 1966 Altitudinal distribution of stoneflies (Plecoptera) in a Rocky Mountain drainage system. Journal of the Kansas Entomological Society 39 4, 668-675.
They refer to this species as Arcynopteryx signata. They never caught any below 8,500 feet and considered Megarcys signata and five other species to be stenothermic or adapted to cold temperatures.
Kondratieff,BC and Baumann,RW 2002 A review of the stoneflies of Colorado with description of a new species of Capnia (Plecoptera: Capniidae). Transactions of American Entomological Society 128 3, 385-401.
Quote from page 397; "Adults are active from April to September, depending on the elevation."
Larson,EI; Poff,NL; Atkinson,CL and Flecker,AS 2018 Extreme flooding decreases stream consumer autochthony by increasing detrital resource availability. Freshwater Biology, 63(12), pp.1483-1497. PDF
Quote: "All taxa showed variability in the proportion of their diet derived from autochthonous compared to allochthonous sources across streams (Supplementary Table 1). Generally, mean consumer autochthony ranged from around 0.25 to 0.5 proportionally of the diet. Certain taxa, such as the rhyacophilid caddisfly Rhyacophila angelita (mean of dietary proportion derived from the epilithon of 0.39-0.48 for their prey across streams), the simuliid blackfly Simulium sp. (0.24-0.31), the perlodid stonefly Megarcys signata (0.48-0.53) and nemourid stonefly Zapada sp. (0.51-0.53) varied little in their resource use across streams. In contrast, the baetid mayfly Baetis bicaudatis (0.22-0.52), the perlodid stonefly Kogotus modestus (0.28-0.61), and the ephemerellid mayflies Drunella doddsi (0.26-0.55), and Drunella grandis (0.25-0.42) exhibited more variability in their resource use across streams. Full stable isotope results and biplots are available in Supplemental Figures 1 and 2."
McIntosh,AR and Peckarsky,BL 1999 Criteria determining behavioural responses to multiple predators by a stream mayfly. Oikos, pp.554-564. PDF
Abstract: "We investigated prey responses to multiple predators by observing drift and positioning of Baetis bicaudatus (Ephemeroptera: Baetidae) nymphs in the presence of non-feeding brok trout (Salvelinus fontinalis chemical cues) and predatory stoneflies (Megarcys signata with glued mouths). In experiments carried out in stream-side channels Baetis antipredator behaviour differed markedly in response to each predator separately; they drifted away from stoneflies, and reduced exposure and drift levels in the presence of trout odour. When both predator cues were presented together the effect of stoneflies, which were observed more frequently at night, depended on the time of day and was modified by presence of fish odour. During the day Baetis responses to both predators were hierarchical as drift and the number on stone tops was reduced whenever fish odour was present regardless of the presence of stoneflies. At night when stoneflies were foraging, Baetis behaviour reflected a compromise between avoiding both predators, with responses to both predators being intermediate between that characteristic of stonefly avoidance (entering the drift) and that characteristic of fish avoidance (drift avoidance). The switch in antipredator behaviour from 'hierarchy' to 'compromise' between multiple predators reflects changes in the relative risk of predation during the day and night. During the day visually feeding trout present a higher predation risk, but at night when stoneflies are foraging and trout visual systems are less effective, the predation risks become more similar. We found no evidence for 'accumulative' avoidance where the magnitude of avoidance increases in response to the number of predators present, because avoiding stoneflies conflicted with avoiding trout. We predict that predator avoidance will take place in the form of a 'hierarchy' when one predator presents substantially higher predation risk than another; 'compromise' behaviour will be observed when the predation risk presented by two predators is more equivalent, but avoiding one predator conflicts with avoiding another; and 'accumulative' antipredatory responses will be observed only when responses to different predators do not conflict. We emphasise that investigation of the responses of prey to multiple predators is important to our understanding of natural communities, and is likely to reveal details about the mechanisms determining the outcome of interactions involving multiple predators."
Molles,MC and Pietruszka,RD 1983 Mechanisms of prey selection by predaceous stoneflies: roles of prey morphology, behavior and predator hunger. Oecologia 57(1) 25-31. Abstract
Needham,JG and Claassen,PW 1925 A Monograph of the Plecoptera of North America. Entomological Society of America, Lafayette, Indiana. 397 pages.
Described as Perlodes signata.








Nelson,CH 2009 Surface ultrastructure and evolution of tarsal attachment structures in Plecoptera (Arthropoda: Hexapoda). Aquatic Insects, (31)523-545. Html
The author used scanning electron microscopy (SEM) to image the plantar surfaces of the stonefly tarsomeres and pretarsus of Megarcys signata and a number of other species.
Obernborfer,RY; McArthur,JV; Barnes,JR and Dixon,J 1984 The effect of invertebrate predators on leaf litter processing in an alpine stream. Ecology, 65(4), pp.1325-1331.
Abstract: " The effect of the predators Megarcys signata (Plecoptera: Perlodidae) and Rhyacophila sp. (Trichoptera: Rhyacophilidae) on the abundance of macroinvertebrates and on the rates of leaf processing in artificial leaf packs was studied by using manipulative field experiments. Predators were confined within artificially constructed leaf packs in an alpine stream in Utah. Both predators significantly reduced the rate of breakdown of leaves in the fall, but had no effect in late winter, when the most important shredder, Zapada cinctipes, emerges, Predation on shredders caused a reduction in breakdown rates and an increase in the residence time of the leaf litter in the stream. In experimental treatments where predators significantly reduced the numbers of shredders and decreased the rate of leaf processing, leaf pack half-life increased an average of 10.3 d. These experiments demonstrate that invertebrate shredders can contribute substantially to the rate of leaf processing in streams and suggest that one of the factors limiting the abundance of natural shredder population is predation."
Peckarsky,BL 1988 Why predaceous stoneflies do not aggregate with their prey. Internationale Vereinigung für Theoretische und Angewandte Limnologie Verhandlungen 23, 2135-2140.
Investigating optimal foraging theory, she found that contrary to theory K. modestus and Megarcys signata larvae did not hang out in high concentrations of prey.
Peckarsky,BL 1990 Habitat selection by stream-dwelling predatory stoneflies. Canadian Journal of Fisheries and Aquatic Sciences 48, 1069-1076.
Peckarsky,BL 1991a A field test of resource depression by predatory stonefly larvae. Oikos 611 3-10.
Peckarsky,BL 1991b Is there a coevolutionary arms race between predators and prey? A case study with stoneflies and mayflies. Advances in Ecology 1, 167-180.
Peckarsky,BL 1991 Mechanisms of intra- and interspecific interference between larval stoneflies. Oecologia 85(4) 521-529.
Abstract: "The purpose of this study was to quantify and compare the behavioral components of the predator-prey interactions between stoneflies and alternative mayfly prey, essential to understanding the reasons for differential rates of predation. This study enables identification of the processes or mechanisms underlying selective predation, and the generation of evolutionary hypotheses to explain why natural selection might favor certain predator behaviors. In situ predation experiments at the East River and Benthette Brook, Gunnison County, Colorado, compared, under near-natural conditions, instantaneous mortality rates due to predation by two species of stream-dwelling stonefly nymphs on different mayfly species. Single-prey experiments predicted relative prey mortality rates in low-density mixed prey assemblages, which we used to test the effects of predator hunger levels and substrate removal on selective predation by stoneflies. One stonefly species, Megarcys signata, consistently selected Baetis bicaudatus over Cinygmula mimus and Ephemerella infrequens in single-prey and mixed-prey trails, when starved for 6 d, 1 d, or fed prior to experiments, and when natural substrates were present or absent. However, substrate removal increased Cinygmula's absolute mortality rate, resulting in significant selectively on this species over Ephemerella. The other stonefly species, Kogotus modestus, selectively consumed either Baetis or Epeorus deceptivus over Cinygmula, depending on the experimental conditions. While Kogotus hunger levels did not affect differential prey mortality, substrate removal resulted in selection of Baetis over Epeorus, and increased absolute mortality rates of all three prey species. These data show that substrate removal can alter patterns of selective predation, which we attribute to differential effects of refuges on predator encounter rates with different prey types. To determine mechanisms underlying observed patterns of selective predation, we videotaped behavioral interactions between predators and prey under simplified field conditions that enabled careful observation. With no substrate present, predator-prey encounter rates and captures per attack were equivalent among prey types (except for Ephemerella, which was never captured). However, both starved and fed predators showed significantly higher probabilities of attack per encounter with Baetis than with any other mayfly species, which resulted from Baetis' hydrodynamically conspicuous swimming motion acting as a proximal attack stimulus. These data suggest that selective predation by stoneflies is more a consequence of active behavioral selection than of prey vulnerability since the probability of attack given an encounter was not constant across all prey types. However, we contend that prey defensive behavior and morphology affect not only encounter rates and capture success, but also attacks per encounter, because prey that did not swim were attacked less per encounter than were prey that swam. Also, although we did not observe significant differences among prey types in encounter rates or captures per attack, a multiple regression analysis showed that these factors, in addition to attacks per encounter, contributed significantly to observed variation in capture rates among prey types in behavioral trials. Thus, we conclude that differential prey vulnerability or fixed behavioral selection is a more probable proximate cause of selective predation than is active behavioral selection, since stoneflies showed a fixed attack response to swimming prey. Finally, we stress that this mechanism of selective predation may or may not result in optimal foraging from an energy maximization standpoint, depending on the direction of competing selection pressures."
Peckarsky,BL 1996 Alternative predator avoidance syndromes of stream-dwelling mayfly larvae. Ecology, 77(6), pp.1888-1905. PDF
Abstract: "Experiments were conducted to compare the patterns, mechanisms, and costs of predator avoidance behavior among larvae of five species of mayflies that co-occur with the predatory stoneflies, Megarcys signata and Kogotus modestus in western Colorado streams. Mayfly drift dispersal behavior, use of high vs. low food (periphyton or detritus) patches, microhabitat use, positioning, and activity periodicity were observed in the presence and absence of predators in circular flow-through chambers using natural stream water. Also, distances from predators at which prey initiated escape responses were compared among prey and predator species. Costs of predator avoidance behavior were assessed by measuring short-term (24 h) feeding rates of mayflies in the presence or absence of predatory stoneflies whose mouthparts were immobilized (glued) to prevent feeding. The intensity and associated costs of predator avoidance behavior of mayfly species were consistent with their relative rates of predation by stoneflies. Megarcys consumes overwintering generation Baetis bicaudatus > Epeorus longimanus > Cinygmula = Ephemerella; Kogotus consumes summer generation Baetis > Epeorus deceptivus = Cinygmula; Megarcys eats more mayflies than Kogotus. While Megarcys induced drift by Baetis, Epeorus, and Cinygmula, this disruptive predator avoidance behavior only reduced food intake by Baetis and Epeorus. The morphologically defended mayfly species, Ephemerella, neither showed escape behavior from Megarcys, nor any cost of its antipredatory posturing behavior. Only Baetis responded by drifting from Kogotus. No mayfly species shifted microhabitats or spent less time on high-food patches in the presence of foraging stoneflies. However, predators enhanced the nocturnal periodicity of Baetis drift, which was negligible in the absence of stoneflies as long as food was abundant. Lack of food also caused some microhabitat and periodicity shifts and increased the magnitude of both day and night drift of Baetis. Thus, Baetis took more risks of predation by visual, drift-feeding fish not only in the presence of predatory stoneflies, but also when food was low or they were hungry. All other mayflies were generally nocturnal in their use of rock surfaces, as long as food was abundant. Finally, the distances at which different mayfly species initiated acute escape responses were also consistent with relative rates of predation. This study demonstrates alternative predator avoidance syndromes by mayfly species ranging from an initial investment in constitutive morphological defenses (e.g., Ephemerella) to induced, energetically costly predator avoidance behaviors (e.g., Baetis). Although the costs of Ephemerella's constitutive defense are unknown, experiments show that prey dispersal is the mechanism underlying fecundity costs of induced responses by Baetis to predators, rather than microhabitat shifts to less favorable resources or temporal changes in foraging activity. A conceptual model suggests that contrasting resource acquisition modes may account for the evolution and maintenance of alternative predator avoidance syndromes along a continuum from Baetis (high mobility) to heptageniids (intermediate mobility) to Ephemerella (low mobility). Prey dispersal (swimming) to avoid capture results in reduction of otherwise high fecundity by Baetis, which trades off morphological defense for enhanced ability to acquire resources. Thus, improved foraging efficiency is the selection pressure maintaining the highly mobile life style in Baetis, which increases resource acquisition and fecundity, offsetting the high mortality costs associated with this behavior."
Peckarsky,BL and Cowan,CA 1995 Microhabitat and activity periodicity of predatory stoneflies and their mayfly prey in a western Colorado stream. Oikos, 513-521. PDF
Abstract: " Experiments were conducted to determine whether overlap between microhabitat preferences and activity periodicities of four mayfly species and their stonefly predators could explain species-specific differences in predator-prey encounter frequencies. Preferences for rock type (slate or granite), flow microhabitat (high or low), rock surface (top, bottom, upstream or downstream sides), and periodicity of drift and the use of rock tops were measured in a stream-side system of flow-through circular Plexiglas chambers receiving natural stream water and light levels. These parameters were compared among the predatory stoneflies, Megarcys signata or Kogotus modestus, and four species of mayflies that vary in their encounter rates with the stoneflies. Based on predator-prey encounter rates previously observed in similar chambers, we expected greater overlap between Megarcys and Ephemerella infrequens and the overwintering generation of the bivoltine mayfly, Baetis bicaudatus than with Cinygmula sp. Likewise, we expected Kogotus microhabitat use to overlap more strongly with that of summer generation Baetis than with later instars of Cinygmula and Epeorus deceptivus. Results ran counter to our predictions, indicating that microhabitats of the prey species with high predator encounter rates did not overlap more strongly with the stoneflies than did mayflies with low predator encounter rates. Most mayflies and stoneflies preferred the bottom surfaces of granite rocks, and showed few flow preferences. Most were nocturnal in their use of top rock surfaces, in drift and feeding activity periodicity. Therefore, nocturnal activity periodicities of both mayflies and stoneflies confirm that mayflies have not evolved feeding periodicity to avoid encounters with foraging stonefly predators. We conclude therefore, that neither temporal nor spatial microhabitat overlap is a reasonable explanation of differential encounter rates between predatory stoneflies and their mayfly prey. Alternative explanations for differential encounter rates are that more abundant or more mobile mayflies have higher encounter rates with predators, and effective pre-contact predator avoidance responses of other mayflies reduce their encounter rates with stoneflies."
Peckarsky, BL; Cowan,CA and Anderson,CR. 1994 Consequences and plasticity of the specialized predatory behavior of stream-dwelling stonefly larvae. Ecology 75(1) 166-181. Abstract
Peckarsky,BL; Cowan,CA; Penton,MA and Anderson,C 1993 Sublethal consequences of stream-dwelling predatory stoneflies on mayfly growth and fecundity. Ecology 74(6):1836-1846. Abstract
Peckarsky,BL; Dodson,SI 1980 Do stonefly predators influence benthic distributions in streams? Ecology 61(6) 1275-1282. Abstract
Peckarsky,BL; Kerans,B; Taylor,BW and McIntosh,AR. 2008 Predator effects on prey population dynamics in open systems. Oecologia.
Rader,RB and Belish,TA 1999 Influence of mild to severe flow alterations on invertebrates in three mountain streams. Regulated Rivers: Research & Management. 15(4)353 - 363.
Discussing Megarcys signata along with three other aquatic insects, they comment that "some stoneflies declined or were even locally extirpated" by severe flow alterations due to dams and water abstraction of their habitat.
Richardson,JW and Gaufin,AR 1971 Food habits of some western stonefly nymphs. Transactions of American Entomological Society 97, 91-121.
Discussed as Arcynopteryx signata, they found that the nymphs emerge April to August. Based on their gut data, Megarcys is mostly carnivorous. They eat mostly Baetidae (small minnow mayflies) and Heptageniidae, as well as lesser amounts of Chironomidae (midges) and Simuliidae (blackflies). A few stoneflies were found in their guts indicating occasional cannibalism or intraguild predation. One specimen´s gut contained strictly plant material, indicating this species can be omnivorous and switch it´s diet when needed.
Smith,LW 1917 Studies of North American Plecoptera (Pteronarcinae and Perlodini) Transactions of the American Entomological Society 43(4):433-489. PDF


Stark,BP and Szczytko,SW 1988. Egg morphology and phylogeny in Arcynopterygini (Plecoptera: Perlodidae) Journal of the Kansas Entomological Society 61(2) 143-160.First Page
Abstract: Comparative data are provided for eggs of nine of the eleven recognized genera in the Holarctic tribe, Arcynopterygini, and these data are used to generate a preliminary phylogeny for the group. Four generic clusters (Megarcys/Sopkalia; Frisonia/Perlinodes/Oroperla; Arcynopteryx/Neofilchneria/Skwala and Setvena/Pseudomegarcys) are established primarily from egg data, but resolution of the Frisonia and Arcynopteryx trichotomies required data from other character suites. Detailed illustrations of the epiproct complex are given for six Nearctic genera to provide additional support for the current generic classification of the group and a standardized terminology is proposed for systellognathan Plecoptera eggs.
Stewart,KW and Kondratieff,BC 2012Larvae of the Nearctic species of the stonefly genus Megarcys Klapálek (Plecoptera: Perlodidae). Illiesia 8(3):16-36. PDF
Abstract: " Associated larvae of the five Nearctic species of Megarcys are comparatively described and illustrated. Four of them, Megarcys irregularis Banks, M. subtruncata Hanson, M. watertoni (Ricker), and M. yosemite (Needham and Claassen) are described in detail for the first time. Body size and setation, wingpad macroptery/brachyptery, and gill size were variable between populations of some species. Diagnostic differences were not found in late instar larvae of the five species in color and pigment patterns, mouthparts, body and appendage setation, gill size and structure, and developing male and female genitalia. The detailed descriptions and measurements help expand the generic diagnosis, show morphological differences between populations of different habitats, and provide an atlas of illustrations for larvae of the five species. "

Look for the orange mites on this female Megarcys nymph. The white nubs among the mites are gills. She was caught and released from the upper East River on the 17th of July 2009. Straight in front of her is a small Cinygmula nymph. There are three Chironomid or Midge larvae around the stonefly as well.
Photo of nymph on page 111 figure 6.43. Illustrations of nymph on page 420-421, figures 14.35-14.36
Taylor,BW; Anderson,CR and Peckarsky,BL 1998 Effects of size at metamorphosis on stonefly fecundity, longevity, and reproductive sucess. Oecologia 114, 494-502. Abstract
Taylor,BW; Anderson,CR and Peckarsky,BL 1999 Egg diapause and semivoltinism in the Nearctic stonefly Megarcys signata (Plecoptera: Perlodidae). Aquatic Insects 21, 179-185.
They found that M. signata was protandrous (males emerge first) and females were about twice the body mass of males in both streams studied. M. signata emerged earlier and larger from a trout stream (East River) than the stoneflies from a smaller fishless tributary (Benthette Brook). Temperature did not affect size at metamorphosis. Many females from Benthette Brook were brachypterous (short-winged), while East River Megarcys females had longer wings. In 1992 East River stoneflies emerged from mid June to mid July with the peak emergence in late June. Benthette brook Megarcys emerged from early July to late August, peaking in Late July. Adults emerged at dawn between 0600 and 088 hours until later in the season when they emerged at night (after 2200 hours). Adults became more active after dark. Manipulative experiments determined that male body mass did not affect reproductive success. Sometimes male drumming attracted females who mated immediately upon finding the male. Other times the first male to stumble on the female mated with her. Mating usually occurred within the first three days after emergence. Copulation generally lasted all night. Females who mated multiple times had lower total lifetime fecundity than females who mated once. Multiple matings reduced the lifespan of males and females. There was no evidence of parthenogenesis in unmated females. Neither sex fed on sugar water except rarely and dissections of field collected adults showed atrophied digestive tracts.
Taylor,BW; Anderson,CR; Peckarsky,BL 1999 Delayed egg hatching and semivoltinism in the Nearctic stonefly Megarcys signata (Plecoptera: Perlodidae). Aquatic Insects, 21(3), pp.179-185. PDF
Abstract: "Delayed egg hatching can influence the survival of aquatic insects by reducing exposure to competitors, predators, parasites, or unfavorable environmental conditions. We examined egg development in a Colorado population of the stonefly, Megarcys signata (Plecoptera: Perlodidae), whose larvae inhabit high altitude streams in the Rocky Mountains of western North America. Five-thousand fertilized eggs were collected from 40 different females and incubated in the East River, Gunnison County, Colorado, until late fall, at which time unhatched eggs were transported to incubation chambers in Ithaca, New York. We used three different combinations of photoperiod and temperature (10L:14D, 4°C; 10L:14D, 8°C; 8L:16D, 8°C) in an attempt to induce hatching. Eggs in the treatments with temperatures elevated above normal winter temperatures in the East River (8°C) began developing after 6 months, whereas eggs in the treatment most closely simulating natural stream conditions (4°C) did not hatch after 10 months of incubation. Our data indicate that this population of Megarcys signata has an extended egg stage that persists for almost a year, and that it is semivoltine at these elevations of 2885 and 2895 meters."
VanWieren,BJ; Kondratieff,BC and Stark,BP 2001 A review of the North American species of Megarcys Klapálek (Plecoptera: Perlodidae). Proceedings of the Entomological Society of Washington 103 2, 409-427.
They provide keys to identify the five North American species by male or females and eggs. There are illustrations of an adult male Megarcys signata on page 410, female subgenital plates on page 418 and four electron micrographs of an egg on page 419. Among the specimens they examined were a male and female collected by B. Armitage from the East River near Avery campground in 1986 and a male and female from tributaries to Oh-Be Joyful Creek in 1996 by B. Kondratieff and N. Lorenzon
Ward,JV, Kondratieff,BC and Zuellig,RE 2002 An Illustrated Guide to the Mountain Stream Insects of Colorado. 2nd ed. University Press of Colorado, Boulder, Colorado. 219 pages.
Illustration of M. signata nymph on page 74, figure 34.
Wellnitz,T 2014 Can current velocity mediate trophic cascades in a mountain stream?. Freshwater Biology, 59(11) 2245-2255. PDF
Zanetell,BA and Peckarsky,B 1996 Stoneflies as ecological engineers-hungry predators reduce fine sediments in stream beds. Freshwater biology, 36(3), pp.569-577.
Abstract: "1. We conducted experiments in a Colorado Rocky Mountain stream to measure the effect of foraging by predatory stoneflies (Megarcys signata) on fine sediment deposition and removal.
2. Cages containing one of four treatments were placed in the stream substratum and removed 3 days after fine sediment load to the stream was experimentally increased. Cages containing a stonefly but no prey accumulated less sediment than cages initially with no benthic invertebrates. Although cages with Megarcys plus prey also tended to have less sediment than controls, sediments were not reduced in cages with prey only.
3. Distance from sediment source, depth and current velocity at cages, final prey density and numbers of prey per predator gut at the end of the experiment had no effects on sediment accumulation in cages.
4. To determine the mechanisms underlying this effect, behavioural tests were conducted in a stream-side artificial stream system. To simulate the different hunger levels of stoneflies in cages, Megarcys were fed or starved for 3 days before behavioural trials that were repeated during high (night) and low (day) stonefly feeding periods. During night trials, foraging movements by starved Megarcys were more prolonged and active than those of fed stoneflies, regardless of the presence of prey. However, neither hunger level nor prey presence affected stonefly foraging behaviour during day trials.
5. Results of field experiments suggest that the presence of stoneflies enhances sediment removal from interstitial spaces. Behavioural observations indicate that nocturnal searching for prey by hungry Megarcys dislodges fine sediments from interstitial spaces.
6. Future studies should explore beyond the direct impacts of predators on stream invertebrate communities, and investigate the potential impact of predators on benthic microhabitat quality.
"
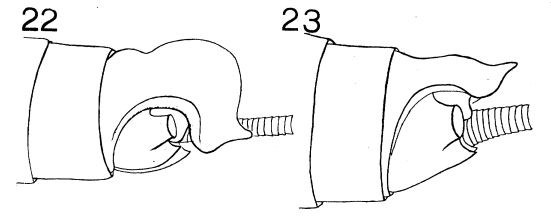
Illustrations

Mesosternal Y-arms.
Only visible easily on mature nymphs. |
|